


Accueil > Nos Actions > Références & Supports > Articles scientifiques et petits essais > Ecosystèmes et biodiversité > Biodiversité 2/5 : Méthodes d'analyses
Introduction
Quelles méthodes d’analyses les chercheurs mettent-ils en place pour mesurer la biodiversité d’un milieu ?
Le Projet
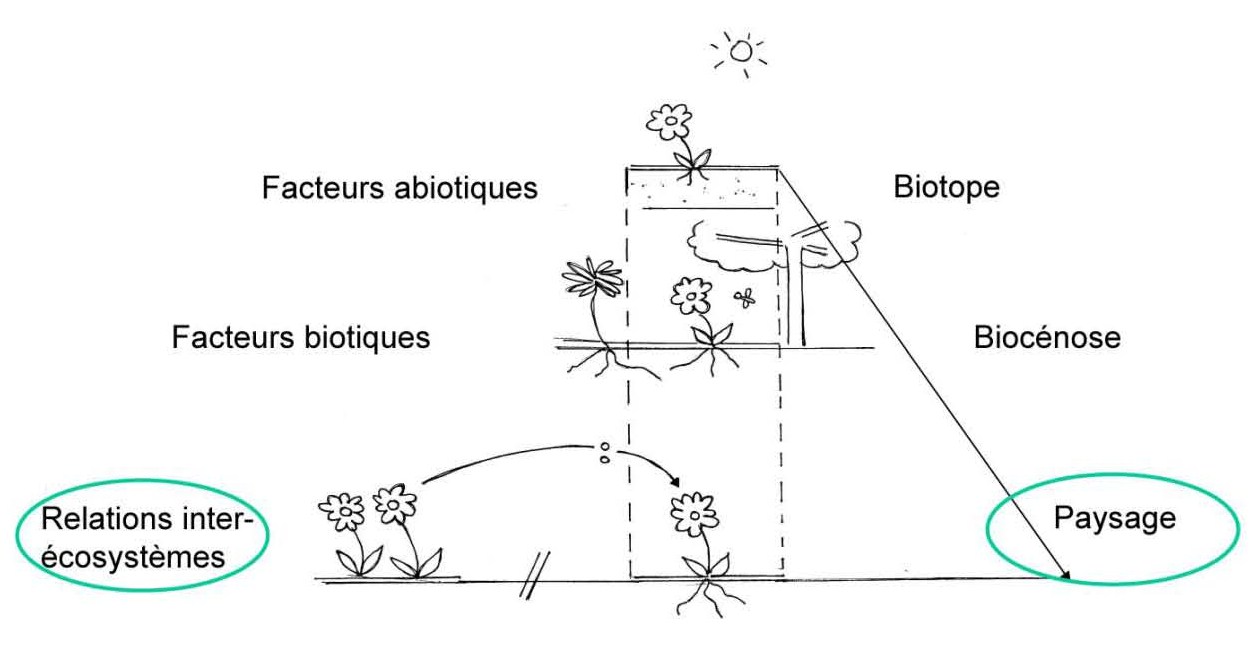
Concept : La biodiversité, c’est des espèces animales, des espèces végétales, les interactions entre ces espèces et avec leurs milieux naturels. Mais quels sont les outils de mesure de tous ces paramètres ?
Échantillonner
Collecte de biote
Au laboratoire, les échantillons prélevés sur le terrain sont normalement triés par espèce selon les guides et les clés standard d’identification relatifs aux groupes taxinomiques et aux zones géographiques des prélèvements.
Dans la plupart des travaux, cette procédure est suivie par un dénombrement des individus par espèce et par la détermination de la biomasse spécifique en poids humide, poids sec et poids sec sans cendre. La mesure de la biomasse autrement qu’en poids humide étant destructive, si des spécimens importants sont présents dans les échantillons prélevés, il faut les retirer avant cette étape. La correction pour la biomasse ainsi perdue est normalement fondée sur les régressions poids humide-poids sec, calculées avec les spécimens qui ne sont pas conservés dans leur intégrité. En présence d’un seul spécimen d’une espèce, il ne faut pas aller au-delà du poids humide pour déterminer la biomasse.
Photographies
Il est plus difficile d’identifier les espèces à partir des photos qu’en examinant les spécimens. En cas de difficulté, des échantillons seront nécessaires pour compléter les photos. Dans de nombreux cas, les photos permettent seulement l’identification jusqu’à un niveau taxinomique assez général. Toutefois, si on apprend à bien connaître l’ensemble complet du biote étudié, des identifications très fiables des espèces sont possibles à partir de gros plans. Pour l’analyse du rapport espèces-superficie, les photos sont tout aussi acceptables que les échantillons.
Évaluer la biodiversité
Les méthodes d’évaluation de la biodiversité doivent avoir deux volets :
![]() l’établissement d’un inventaire des espèces
l’établissement d’un inventaire des espèces
![]() la compréhension des changements de la structure des communautés dans le temps et dans l’espace, c’est-à-dire les différences parmi les assemblages d’espèces.
la compréhension des changements de la structure des communautés dans le temps et dans l’espace, c’est-à-dire les différences parmi les assemblages d’espèces.
Pour simplifier les vastes ensembles de données obtenus, on emploie plusieurs techniques, qui entrent dans l’une des cinq grandes rubriques décrites ci-dessous. Pour toutes ces procédures, divers traitements préalables des données sont nécessaires avant l’essai d’une structure. Clarke et Green (1988) ont résumé ces étapes ainsi que d’autres aspects des plans statistiques.
1. Méthodes uni variées
Ces méthodes servent habituellement à faire ressortir les caractéristiques générales des communautés qui ne sont pas fonction de taxons spécifiques ; elles sont donc indépendantes de l’espèce et ne sont pas sensibles aux variations spatio-temporelles dans la composition spécifique, de sorte que les assemblages n’ayant pas d’espèces en commun peuvent théoriquement présenter des diversités égales. Elles sont plus faciles à utiliser que les méthodes multi variées mais, tout comme les méthodes graphiques et distributionnelles, elles ne sont pas aussi sensibles pour détecter les changements (Warwick et Clarke, 1991).
a) Richesse spécifique
a) Richesse spécifique
Il s’agit de la mesure de la diversité la plus ancienne et la plus élémentaire, fondée directement sur le nombre total d’espèces dans un site ; on préfère l’expression « richesse spécifique », car le nombre exact d’espèces dans une communauté est rarement connu.
Toutefois, cette méthode dépend de la taille des échantillons et ne considère pas l’abondance relative des différentes espèces. Sa valeur écologique est donc limitée. En tant que concept écologique, l’abondance est une autre composante importante de la diversité (Hurlbert, 1971), que Peet (1974) définit comme l’hétérogénéité, c’est-à-dire l’équitabilité ou l’égalité de la répartition des individus parmi les espèces. Un grand nombre d’espèces fait augmenter la diversité spécifique, et une distribution égale ou équitable parmi les espèces représente aussi une plus grande diversité. On a créé plusieurs indices à cet égard, mais il est inutile de tous les calculer, car ils sont fortement corrélés (Gray et al., 1992). L’indice de Shannon-Wiener est le plus utilisé.
b) Indice de Shannon-Wiener
b) Indice de Shannon-Wiener
Avec pi = abondance proportionnelle ou pourcentage d’importance de l’espèce, se calcule ainsi : pi = ni/N ; S = nombre total d’espèces ;
ni = nombre d’individus d’une espèce dans l’échantillon ;
N = nombre total d’individus de toutes les espèces dans l’échantillon.
Ainsi, la valeur de H’ dépend du nombre d’espèces présentes, de leurs proportions relatives, de la taille de l’échantillon (N) et de la base de logarithme. Le choix de la base de logarithme est arbitraire (Valiela, 1995) mais, lorsqu’on compare des indices, la base utilisée doit être mentionnée et être la même.
La dominance marquée d’une espèce révèle une faible diversité, alors que la co-dominance de plusieurs espèces révèle une grande diversité. Puisque l’équation est un estimateur biaisé (Valiela, 1995).
Un terme correctif (S-1)/2N doit être soustrait du membre droit de l’équation.
Un essai formel des différences statistiques dans les valeurs de H’ est possible en calculant l’écart de H’ (Hutcheson, 1970). Štirn décrit des calculs pratiques pour appliquer cet indice à des données ( 1981). De tous les indices, la formule de Shannon et de Wiener est probablement l’indice le plus utilisé qui considère à la fois l’abondance et la richesse spécifique (Gray et al., 1990). Elle semble constituer le moyen le plus utile pour obtenir des indices de diversité significatifs, relativement indépendants de la taille des échantillons (Štirn, 1981). Certains (Štirn, 1981 ; Gray et al., 1992) l’ont recommandée comme étant la meilleure expression de la diversité du biote.
L’équitabilité
C’est le rapport de la diversité observée à une distribution de fréquence des espèces complètement égale (échelle de 0-1), peut être quantifiée séparément à l’aide de l’indice Shannon-Wiener comme étant :
| J’ = H’/H’max |
où H’ est la diversité spécifique observée et H’max est le logarithme du nombre total d’espèces (S) dans l’échantillon (ACMRR/IABO, 1976 ; Gray et al., 1992).
Par exemple, deux espèces présentant 50 individus chacune représenteraient une équitabilité ou une égalité complète d’une valeur de 1. Deux espèces, dont la première est représentée par un individu, et la deuxième, par 99, obtiendraient seulement un résultat de 0,08.
2. Méthodes graphiques et distributionnelles
Ces méthodes font partie d’une classe de techniques située à mi-chemin entre les sommaires uni variés et les analyses multi variées complètes de la matrice espèces/échantillons. Deux méthodes sont couramment utilisées pour comparer la diversité du biote.
a) Courbes de raréfaction de la diversité
a) Courbes de raréfaction de la diversité
Cette méthode, élaborée par Sanders (1968), est largement utilisée pour évaluer la diversité au sein des écosystèmes, par exemple dans les études sur la pollution (Gray et Pearson, 1982). Elle estime le nombre d’espèces pour un nombre donné d’individus. Son principal avantage est qu’elle ne dépend pas de la taille des échantillons. Elle prend pour hypothèse que les rapports espèces/individus sont semblables dans les communautés comparées et que les individus sont distribués aléatoirement. Štirn (1981) et Sanders (1968) montrent comment construire les graphiques.
b) Courbes de dominance k
b) Courbes de dominance k
Mises au point par Lambshead et al. (1983), ces courbes sont obtenues en traçant un graphique du pourcentage de l’abondance cumulative par rapport au rang k de l’espèce sur une échelle logarithmique, où l’assemblage d’espèces x est plus divers que y si la courbe de y est toujours au-dessous de celle de x ou la touche (voir Warwick, 1986 ; Warwick et al., 1990a,b). Clarke (1990) a élaboré une procédure statistique pour tester les différences significatives par « analyse des similarités » (ANISE), en comparant la variabilité des courbes de dominance k d’échantillons répétés à celle d’échantillons séparés spatialement ou temporellement.
On peut également superposer des courbes de dominance k représentant l’abondance et la biomasse des espèces (Warwick, 1986). On appelle ces courbes superposées « courbes de comparaison abondance-biomasse » (ABC) ; la position relative de l’abondance et de la biomasse sur le graphique peut révéler les impacts de la pollution. Pour les sites relativement peu perturbés, les courbes de biomasse se trouvent au-dessus des courbes d’abondance, et vice versa. Cette méthode est très répandue (Warwick et Clark 1991 ; Clark, 1990 ; Warwick, 1988) mais, pour la mission du RESE, elle n’a qu’une applicabilité limitée, car les données sur la biomasse ne sont et ne seront probablement pas recueillies sur une base régulière.
En général, les deux méthodes (raréfaction et dominance k) donnent les mêmes résultats. Les courbes de dominance k présentent deux avantages : la dominance relative des espèces les plus courantes ou les plus rares peut être déterminée en un coup d’œil, et les calculs permettant de tracer les courbes sont moins compliqués (Lambshead et al., 1983). Comme l’affirment Lambshead et al. (1983), il est recommandé d’appliquer routinièrement les méthodes graphiques aux données de la biologie marine avant de calculer les indices compliqués de diversité ou d’équitabilité.
3. Méthodes multi variées
Dans un contexte écologique, ces méthodes informatisées de tri sont utilisées pour classer par groupes les taxons ou les sites montrant des attributs semblables. Au départ, elles indiquent simplement le degré de similarité ou de dissemblance dans la composition des espèces entre des stations ou à l’intérieur de la même station au fil du temps.
On ne peut obtenir des preuves de corrélation solides des causes et des effets, par exemple ceux des polluants, qu’en reliant les groupements de stations aux gradients mesurés de l’environnement et de la pollution ou à des mesures indirectes de l’intensité de la pollution, notamment la distance par rapport à la source de pollution ou le temps écoulé depuis l’événement polluant (Gray et Pearson, 1982 ; Warwick, 1987).
Parce qu’elles sont fondées sur des critères formels, ces méthodes semblent plus objectives que d’autres. Contrairement aux indices de diversité, les méthodes multi variées décrites préservent l’identité des espèces et sont généralement considérées comme plus sensibles pour la détection des patrons changeants dans les communautés. Elles permettent de détecter les effets plus rapidement (Warwick et Clarke, 1991 ; Gray et al., 1990).
Les méthodes multi variées, toutefois, ont des lacunes. Elles sont considérablement plus complexes que les autres méthodes et nécessitent des traitements préalables substantiels ou une préparation des données, par exemple des transformations, mais il n’existe actuellement aucune procédure uniforme ou acceptée. La matrice de données doit également être réduite en vue du traitement des données. Généralement, cette réduction élimine de l’analyse les espèces rares, qui sont pourtant une propriété intrinsèque de toutes les communautés et peuvent constituer certaines des espèces déterminantes (Gray et Pearson, 1982). Les méthodes multi variées entrent dans les deux grandes catégories du regroupement et de l’ordination. Avant de soumettre les matrices de données à l’un de ces types d’analyse, les données sur l’abondance et la biomasse des espèces sont soumises à une transformation, et on calcule les similarités entre les paires d’échantillons à l’aide du coefficient Bray-Curtis (Bray et Curtis, 1957) : Cz = 2w/(a+b), où a est la somme des abondances de toutes les espèces trouvées dans un échantillon donné ; b, la somme des abondances des espèces d’un autre échantillon ; w, la somme des valeurs d’abondance les plus faibles pour chaque espèce commune aux deux échantillons.
Méthodes de classification
Ces méthodes sont fondées sur l’attribution d’entités aux classes ou aux groupes, les données d’entrée étant généralement les abondances des espèces dans une matrice croisée échantillons/espèces (Gauch, 1982). Le processus de classification est essentiellement une réduction de l’information, pour chaque échantillon, d’un grand nombre de chiffres (toutes les abondances des espèces) en un seul chiffre (attribution du regroupement). Les méthodes de réduction du nombre de chiffres sont innombrables ; elles peuvent mettre l’accent sur les espèces dominantes, les espèces mineures, les espèces individuelles, etc. La méthode la plus couramment retenue effectue le regroupement hiérarchique, les stations semblables étant fusionnées pour former des groupes de plus en plus gros. Ce regroupement est basé sur l’analyse des groupes ou sur le tri à liens simples d’une matrice de similarités entre échantillons, à l’aide de la mesure de similarité de Bray-Curtis. Les résultats sont affichés dans un dendrogramme. On repère les espèces qui contribuent le plus à la division des sites en groupes grâce au programme des pourcentages de similarité (SIMPER) (Warwick et al., 1990a).
Méthodes d’ordination
Ces méthodes donnent un aperçu des relations de similarité entre échantillons en termes d’abondance ou de biomasse des espèces, la distance relative d’une paire d’échantillons reflétant leur dissemblance relative. Clark et Green (1988) les définissent comme une analyse d’une matrice de données de n échantillons par p espèces, grâce à laquelle on obtient une nouvelle série de variables qui prédisent de façon optimale la structure des relations entre les variables originales p. Les méthodes diffèrent selon le critère d’optimalité et selon la façon dont l’algorithme d’ordination détermine les nouveaux axes représentant les nouvelles variables. Actuellement, il existe plusieurs techniques d’ordination, dont l’analyse factorielle des correspondances (AFC) et l’analyse factorielle des correspondances redressée (DECORANA). Warwick (1987) a conclu que le choix de la technique la plus appropriée dépend grandement des préférences personnelles et de la disponibilité des programmes adaptés et des installations informatisées.
Dans les ACP, l’étendue de l’écart représenté par les nouveaux axes est maximisée, car elle est déterminée par une analyse des valeurs et vecteurs propres sur la matrice de corrélation p par p, les nouveaux axes n’étant pas corrélés. La procédure est relativement simple, mais les nouveaux axes peuvent rarement être interprétés comme des facteurs environnementaux responsables de la structure des données sur l’abondance des espèces.
L’analyse multidimensionnelle construit une carte des sites, dans laquelle plus deux échantillons se ressemblent en termes d’abondance ou de biomasse, plus ils sont proches l’un de l’autre sur la carte (Clarke et Green, 1988 ; Gray et al., 1988). Ce type d’analyse est répandu, car il dépend seulement des renseignements de corrélation plutôt que des valeurs quantitatives, et utilise des affirmations telles que « l’échantillon 1 ressemble davantage à l’échantillon 2 qu’à l’échantillon 3 ». La mesure dans laquelle ces rapports peuvent être adéquatement représentés sur une carte bidimensionnelle est exprimée comme statistique de coefficient de stress, les valeurs faibles indiquant le taux de réussite (p. ex. < 0,1). Les résultats sont affichés dans des graphiques dont la configuration et l’échelle sont arbitraires.
Les méthodes de regroupement et d’ordination ne sont pas en concurrence (Clark et Green, 1988) ; on les recommande toutes les deux (Gray et al., 1988).
4. Espèces indicatrices
L’examen des espèces indicatrices effectué par Pearson et Rosenberg (1978) a montré que les espèces présentes dans les endroits les plus pollués, par exemple le polychète Capitella capitata, sont typiques des premiers stades de succession. On peut également trouver ces espèces en grandes densités dans des endroits autres que ceux qui présentent un enrichissement organique (Gray et Pearson, 1982). Néanmoins, des groupes d’espèces caractérisant divers stades d’enrichissement se retrouvent dans des zones peu étendues. Par contre, comme ces groupes varient selon la région géographique, ils ne constituent peut-être pas des indicateurs universels.
Gray et Pearson (1982) ont utilisé la distribution des individus au sein d’une espèce pour identifier les taxons critiques. Contrairement aux méthodes multi variées, il s’agit d’une technique simple, utilisant l’ensemble des données sur les espèces communes et rares, qui identifie les groupes d’espèces indicatrices pouvant servir dans les programmes de surveillance à petite échelle. On porte le nombre d’espèces sur l’axe des ordonnées, et le nombre d’individus par espèce agrégé en classes géométriques, sur l’axe des abscisses. Les courbes temporelles ou spatiales peuvent ensuite être comparées, et on note les changements dans le nombre d’espèces dans une classe de taille particulière. Les perturbations environnementales sont indiquées par une diminution des espèces rares les plus proches de l’axe des ordonnées. Les espèces indicatrices se retrouvent dans les catégories qui comprennent les groupes d’espèces assez courantes (habituellement V et VI) chez lesquelles on observe des changements rapides le long d’un gradient spatial ou temporel (pour de plus amples détails, voir Gray et Pearson, 1982, p. 116-117 ; Gray et al., 1990, p. 290). À l’aide de l’analyse de variance standard (ANOVA), on peut tester les changements d’une espèce particulière pour en vérifier la signification statistique (Gray et al., 1990 ).
Dans le cas des techniques multi variées, on peut aussi identifier les espèces indicatrices qui contribuent le plus à la division des sites en regroupements en utilisant les pourcentages de similarité (SIMPER), comme le décrivent Warwick et al. (1990a).
5. Réductions taxinomiques : agrégation des données sur les espèces à des échelons taxinomiques plus élevés
Par l’analyse de groupement et l’ordination multidimensionnelle, Gray et al. (1990) ont démontré qu’on peut nettement réduire les coûts de surveillance de l’environnement en travaillant à des échelons taxinomiques supérieurs à celui de l’espèce. Quand on ordonne les données sur les espèces en familles, le groupement des sites non pollués, quelque peu pollués et fortement pollués demeure intact et comparable à l’analyse faite au niveau de l’espèce. Avec quelques changements mineurs, le patron général peut rester le même si on regroupe les données en taxons supérieurs : polychètes, mollusques, échinodermes et crustacés. En utilisant l’analyse multidimensionnelle et les graphiques ABC au niveau de la famille, Warwick (1988) n’a trouvé aucune perte d’information, contrairement aux analyses faites au niveau de l’espèce.
Il y avait cependant quelques différences quand on utilisait l’analyse multidimensionnelle au niveau du phylum, selon la force de la transformation, et selon qu’on considérait l’abondance ou la biomasse. Warwick et al. (1990b) ont également trouvé une certaine perte d’information au niveau du phylum ; par contre, pour la macrofaune, la perte était faible avec le groupement des données sur les espèces au niveau de la famille, que ce soit d’après l’analyse uni variée (graphiques ABC) ou l’analyse multi variée (analyse multidimensionnelle). Toutefois, pour les nématodes de la méiofaune, il y avait une perte importante d’information au-dessus du niveau du genre. Warwick et al. (1990b) pensent que le niveau du genre peut être le niveau taxinomique optimal pour l’efficacité de la discrimination, mais que le niveau de la famille peut être approprié pour la macrofaune (Gray et al., 1992).
Seuil
Dépasser un certain seuil peut provoquer des changements rapides et considérables en termes de biodiversité et de bienfaits que l’écosystème peut procurer aux êtres humains. Ce phénomène a été observé dans les écosystèmes aquatiques ouverts, après qu’un seuil de température a été franchi, ou quand les ressources étaient surexploitées. Par exemple, un apport accru de nutriments peut transformer des récifs riches en corail en un écosystème dominé par les algues, moins diversifié et productif d’un point de vue biologique. Des espèces envahissantes peuvent également déclencher des changements spectaculaires dans la structure d’un écosystème. Par exemple, l’introduction d’un animal carnivore semblable à la méduse dans la Mer Noire a engendré la disparition de 26 espèces de premier plan pour la pêche et a contribué à la croissance de la zone « morte » dépourvue d’oxygène.
Témoignages
Suivez notre actualité sur les réseaux sociaux :

{kind=link}