

Accueil > Nos Actions > Références & Supports > Articles scientifiques et petits essais > Evolution > Evolution de l'interaction insecte-plante-champignon sur des gradients d'altitudes.
Introduction
Une des questions fondamentales en écologie est de comprendre les différents facteurs qui mènent à des variations au sein d’interaction d’espèces dans leur habitat. Dans le but de répondre à cette question, nous pouvons utiliser la relation insecte - plante - champignon. Le fait d’y ajouter un facteur tel que l’altitude revient à modifier considérablement cette relation. Dans quelle mesure l’altitude peut modifier cette interaction tripartite ? Autrement dit, quelles sont les effets de l’altitude sur les insectes, plantes et champignons ? Est-ce l’unique facteur faisant varier cette interaction ?
Le Projet
Mycorhize et défense des plantes
Les mycorhizes
Sur terre, environ 80% des plantes vasculaires ont une relation symbiotique avec un champignons (Koricheva et al. 2009). Cette relation est aussi appelée “mycorhize”. Elle est dites symbiotique car chacun des organismes en tire un bénéfice essentiel à leur survie et qui se traduit par des échanges de nutriments. Elles se regroupent sous deux grands types : ectomycorhyzes et endomycorhyzes. Ce dernier étant encore divisé en trois sous-groupes. Nous nous intéresseront ici endomycorhyzes à arbuscules qui représente la majeure partie des symbioses mycorhyziennes sur la planète.

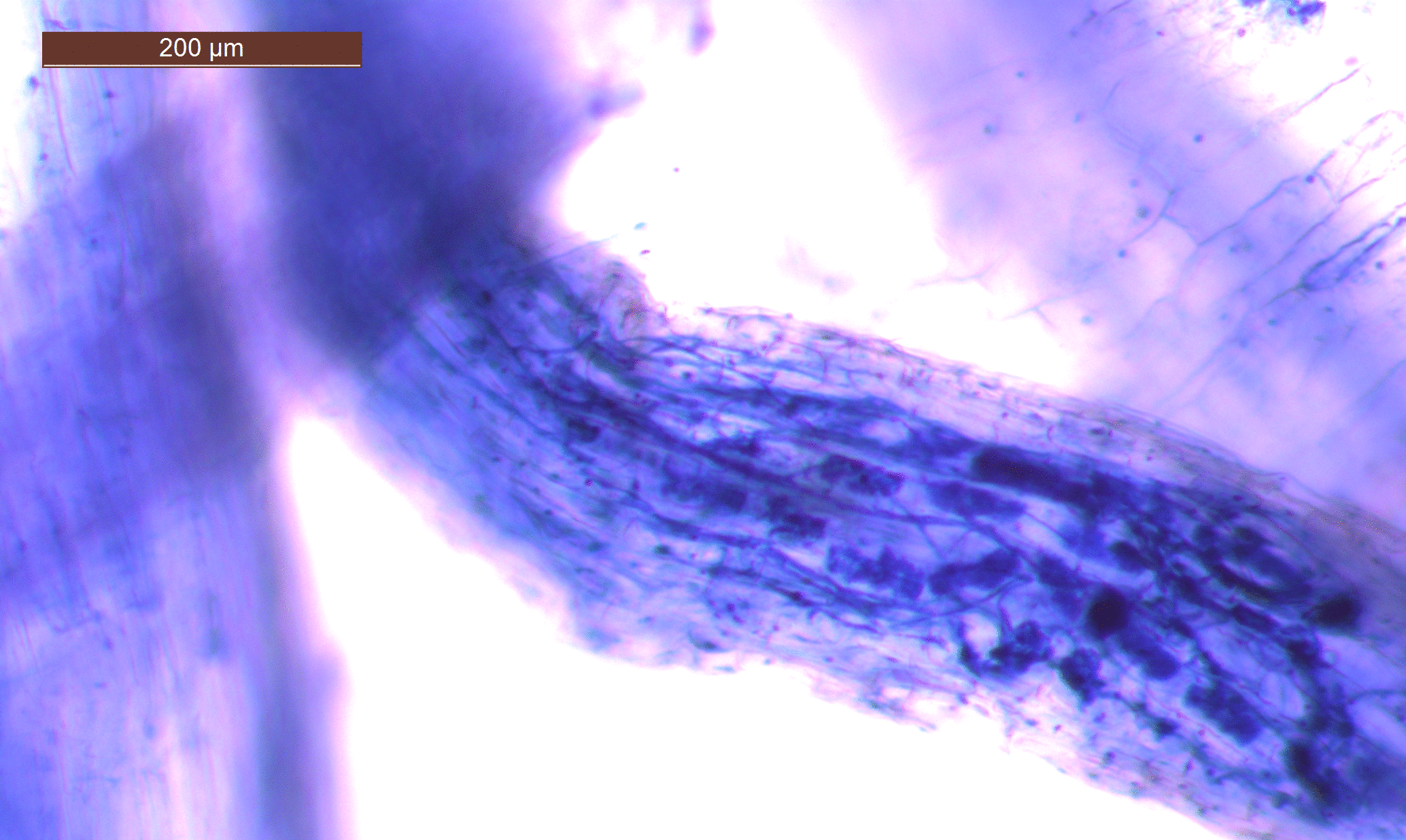
- Mycorhizes arbusculaires dans une racine de Plantain lancéolé
- Photo faite à la loupe binoculaire. On y voit un fragment de racine de Plantain lancéolé. Les compartiments représentent les cellules, Les structures foncées à l’intérieur sont les arbuscules formées par le champignons. C’est ici que se font les échanges entre plante et champignon.
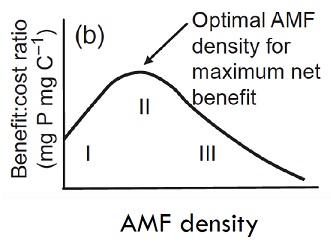
Pour plus de détail, il est possible de se référer à l’article de Mr. Olivier Lavaud (http://www.osi-perception.org/Prese...). Dans le cas des mycorhizes à arbuscules, la symbiose n’est pas tout le temps bénéfique et dépend de la densité du réseau mycélien. Cela peut se traduire par la notion de ratio : coût/bénéfice (Vannette et Hunter. 2011). Lorsque le champignon commence à se développer et coloniser la plante, la densité du réseau est trop faible pour que la dite plante obtienne des nutriments. A l’inverse, plus le champignon s’étend et plus les coûts pour la plante pour maintenir la relation sont important. Dans ce dernier cas le champignon agit comme un parasite. Les deux partis tirent donc bénéfice de cette relation à un stage intermédiaire de développement du réseau mycélien. Le champignon agit ici comme un mutualiste.
La défense des plantes
Interviennent ensuite les insectes herbivores qui se nourrissent des plantes. Ceux-ci survivent en se nourrissant des parties aériennes de la plante, en perçant les tissus afin de capter la sève ou en attaquant les parties souterraines. Il est évident que lorsque une plante est attaquée par un insectes, elle ne se laisse pas faire. Elle met en place tout un système de défense pour y faire face. Ce mécanisme de défense peut se résumer en trois catégories : tolérance, résistance constitutive et résistance induite (Bennett et al. 2006). La tolérance se réfère à la capacité de la plante à surmonter les dommages par compensation des tissus perdus en produisant des structures végétatives ou en induisant des mécanismes de reproduction. La résistance constitutive inclut tous les mécanismes « pré-activés » dans le but d’anticiper toute éventuelle attaque. La résistance induite se définit par la capacité d’une plante à acquérir des adaptations environnementales après une attaque dans le cas où elle devrait à nouveau être confrontée à une agression du même type (Karban et Baldwin. 1997). Il est à préciser ici que la plante peut exprimer ces trois réponses en même temps ou bien une des trois en fonction du type et de l’intensité de l’attaque (Bennett et al. 2006). L’application de ces trois réponses dépend de plus des facteurs biotiques (relation de la plante avec les autres organismes et microorganismes) et abiotiques (par exemple l’environnement).
L’altitude comme variable environnementale
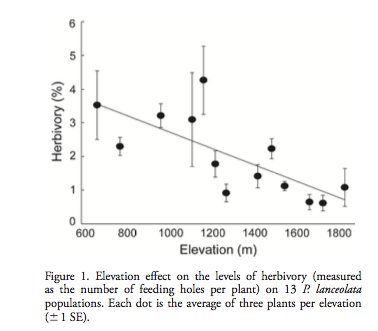
L’altitude est un facteur essentiel de variation. Par analogie, si nous regardons à l’échelle du globe et en se basant au niveau de l’équateur, il y a plus de stabilité environnementale. Cela se traduit par une des interactions biotiques plus importante, une pression plus élevée de la part des herbivores et une production des sols en terme de nutriments plus élevées (Coley et Barone. 1996). L’inverse se produit en se plaçant à des latitudes plus élevées. Les interactions biotiques diminuent tout comme la pression exercée par les herbivores ainsi que la production des sols (Coley et Barone. 1996). Une étude a de plus montré que le nombre d’insectes herbivores était fonction de l’altitude (Pellissier et al. 2014). Nous comprenons donc que l’altitude devient un facteur fondamental de la variation puisqu’il agit sur la température. Cette dernière affectant particulièrement le métabolisme primaire (ce qui attrait à la vie) et secondaire (ce qui attrait aux défenses) de la plante (Pellissier et al. 2014).

- Différence de croissance entre basse et haute et altitude.
- Cette photo met en évidence la différence de production de biomasse aérienne entre une plante de basse altitude (à gauche) et une plante de haute altitude (à droite). Le conditions de basse altitude étant plus favorable (température plus élevée) à la croissance, la plante de gauche possède une biomasse aérienne nettement plus importante.
A basse altitude
Au niveau de la plaine (étage collinéen, environ 500 m), les conditions environnementale sont plus stables et la température y est plus élevée qu’en montagne. Cela induit pression des insectes herbivores plus élevées qui se traduit par le fait que le nombre d’espèce présente et le nombre d’individu par espèce sont plus grands. Cette température favorise aussi une meilleure production au niveau du sol, ce qui favorise la croissance des plantes. Ces dernières étant plus grandes, elles offrent une pus grande surface d’attaque aux insectes. La production du sol favorisée par la température permet aussi d’augmenter le taux de colonisation de la plante par les mycorhizes. La croissance de la plante sera donc d’autant plus favorisée, elle va donc axer son métabolisme sur un caractère plutôt défensif étant donné la pression exercée par les insectes herbivores.
A haute altitude
En montagne (étage montagnard-subalpin, environ 1500 m), les conditions environnementales plus instables et la température moins élevées entraînent une pression des insectes herbivores moins élevées. Il y a donc moins d’espèces représentées et moins d’individus présents (Rasmann et al. 2014). Cette température implique aussi un ralentissement de l’écosystème sol qui est alors nettement moins productif (Hodkinson. 2005). Les plantes ont une croissance plus faible qu’en plaine et sont moins colonisées par les champignons mycorhiziens. Etant donné que leur croissance est défavorisée et qu’elles sont moins attaquées, les plantes vont diriger leur métabolisme vers un axe de croissance des tissus au détriment de la défense.
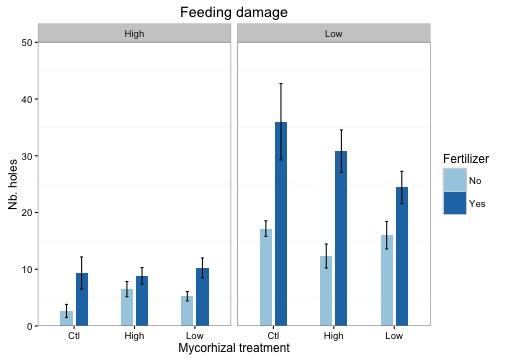
- Dommages des plantes en fonction de l’altitude et du type de mycorhize.
- Les dommages infligés aux plantes par les insectes se traduit par le comptage des trous sur les feuilles. Ce graphe se divise en deux. A gauche les plantes de haute altitude et à droite celle de basse altitude. Dans les deux cas il y a 3 traitements mycorhiziens : contrôle = pas de mycorhizes, high = mycorhize de haute altitude et low = mycorhize de basse altitude. Les barres foncées représentent les plantes ayant reçues du fertilisant et les claires celles qui n’ont rien reçues (traitement additionnelle pour l’expérience).
Ce graphe met en évidence les dommages infligés par des insectes sur des plantes exposées à haute (Col de Chaude et Morcles) et basse altitude (Noville et Lavey). Nous voyons très nettement que les plantes de basse altitude sont beaucoup plus attaquées, ce qui rejoint l’idée exposée par Pellissier. Le nombre d’insectes herbivores est fonction de l’altitude. A basse altitude, il fait plus chaud. Cela permet une plus grande représentation d’espèce et plus d’individus par espèce pouvant attaquer les plantes de manières plus importante.
Un autre effet mis en évidence ici est celui du fertilisant. Systématiquement, dès que les plantes ont reçu de l’engrais, elles subissent plus de dommages. Cela peut se traduire par le fait qu’elles produisent plus de biomasse aérienne et donc plus de surface exposable aux insectes qui ont plus à manger. Nous remarquons enfin que pour les plantes de basse altitude, celles-ci sont moins attaquées lorsqu’elles reçoivent des mycorhizes de basse altitude. Cela peut s’expliquer par une adaptation génétique préférentielle entre plantes et mycorhizes du bas (voir chapitre de génétique plus bas). Cela peut aussi s’expliquer par le fait que le sol du bas étant plus productif, les mycorhizes vont aider les plantes à mieux se défendre puisqu’elles subissent une pression plus importante de la part des insectes herbivores.

- Biomasse aérienne produite (g) en fonction de l’altitude et du type de mycorhize.
- La croissance des plantes se traduit par la biomasse aérienne produite : feuilles et tiges. Ce graphe se divise en deux. A gauche les plantes de haute altitude et à droite celle de basse altitude. Dans les deux cas il y a 3 traitements mycorhiziens : contrôle = pas de mycorhizes, high = mycorhize de haute altitude et low = mycorhize de basse altitude. Les barres foncées représentent les plantes ayant reçues du fertilisant et les claires celles qui n’ont rien reçues (traitement additionnelle pour l’expérience).
Le graphique ci-dessus met en évidence l’effet de la température à haute (Col de Chaude et Morcles) et basse altitude (Noville et Lavey). Comme expliqué précédemment, à basse altitude, il fait plus chaud et les sols sont plus productifs donc les plantes ont une croissance plus importante. Nous voyons que la biomasse aérienne produite est nettement plus élevée pour les plantes de basse altitude (à droite sur le graphique). Ce graphique met aussi en évidence une adaptation des plantes avec leurs mycorhizes (rubrique génétique, voir plus bas). Nous pouvons constater que les plantes de basse altitude ayant reçue des mycorhizes de basse altitude produisent plus de biomasse aérienne que des plantes de basse altitude ayant reçue des mycorhizes de haute altitude. Le même raisonnement peut être appliqué pour les plantes de haute altitude. Cela signifie que même si les mycorhizes de haute altitude devaient favorisées la croissance de la plante, il y a un effet non négligeable de la génétique.
Et la génétique ?
Nous voyons ici qu’un paramètre peut engendrer énormément de variation. Dans la nature il est évident que la température n’est pas le seul facteur à influer sur cette relation tripartite. Une étude a effectivement mis en évidence que la forte variation dans l’interaction insecte-plante-champignon pourrait être due à la variation génotypique des champignons (Li et al. 2014 & Ehinger et al. 2009). En d’autre terme cela signifierai que des espèces de champignon sont adaptés à une certaine espèce de plante et pourrait créer des mycorhizes préférentielles. De plus, si des plantes sont adaptées à l’altitude, il en va de même pour les champignons.
A basse altitude, étant donné que la plante a plus de ressources disponibles et plus de pression de la part des insectes herbivores, elle axe son métabolisme sur la défense. Le champignon peut s’adapter au besoin de la plante et donc indirectement à son environnement en produisant des substances défensives.
A haute altitude, c’est l’inverse. la plante axe son métabolisme sur la croissance car moins de ressources à disposition et moins de pression des insectes herbivores. Le champignon va s’adapter à cette situation en créant un réseau mycélien plus dense dans le sol dans le but d’obtenir plus nutriments.
Pour aller plus loin
Nous ne mentionnons ici que deux facteurs pouvant faire varier drastiquement cette interaction. Comme mentionné plus haut, il est évident que dans la nature, il faut tenir compte de beaucoup plus d’éléments. Nous pouvons mentionner par exemple la structure formée par le champignon au sein des racines de la plante qui peut influer les échanges. Enfin un facteur essentiel dans la modification de cette interaction est l’humain. Nous exerçons une pression démographique non négligeable sur notre environnement qui se traduit par une diminution des espèces présente sur terre. Cette disparition se fait aussi ressentir au niveau des champignons dont certaines variétés ont déjà disparue (Zubek et al. 2016).
Références
Bennett, A. E., Alers-Garcia, J., & Bever, J. D. (2006). Three-way interactions among mutualistic mycorrhizal fungi, plants, and plant enemies : hypotheses and synthesis. The American Naturalist, 167(2), 141–152.
Ehinger, M., Koch, A. M., & Sanders, I. R. (2009). Changes in arbuscular mycorrhizal fungal phenotypes and genotypes in response to plant species identity and phosphorus concentration. The New Phytologist, 184(2), 412–423.
Hodkinson, I. D. (2005). Terrestrial insects along elevation gradients : species and community reponses to altitude. Biological reviews, 80(03), 489-513.
Karban, R., and I. T. Baldwin. 1997. Induced responses to herbivory : interspecific interactions. University of Chicago Press, Chicago.
Koricheva, J., Gange, A. C., & Jones, T. (2009). Effects of mycorrhizal fungi on insect herbivores : A meta-analysis. Ecology, 90(8), 2088–2097.
Li, X., Gai, J., Cai, X., & Li, X. (2014). Molecular diversity of arbuscular mycorrhizal fungi associated with two co-occurring perennial plant species on a Tibetan altitudinal gradient, 95–107.
Pellissier, L., Roger, A., Bilat, J., & Rasmann, S. (2014). High elevation Plantago lanceolata plants are less resistant to herbivory than their low elevation conspecifics : Is it just temperature ? Ecography, 37(March), 1–10.
Rasmann, S., Alvarez, N., Pellissier, L. (2014). The Altitudinal Niche-Breadth Hypothesis In Insect-Plant Interactions. Annual Plant Reviews, 47, 339-360
Vannette, R. L., & Hunter, M. D. (2011). Plant defence theory re-examined : nonlinear expectations based on the costs and benefits of resource mutualisms, 66–76.
Zubek, S., Nobis, M., Błaszkowski, J., Nowak, A., Majewska, M. L., Bąba, W., … Bąba, W. (2016). all aspects of plant biology arbuscular mycorrhiza and fungal root endophytes of weeds in an altitudinal gradient in the Pamir Alai Mountains of Central Asia, 3504(February).
Témoignages
Suivez notre actualité sur les réseaux sociaux :

{kind=link}